New Phytologist
TCP10 acts downstream of light and retrograde signaling pathways to promote Arabidopsis cotyledon de-etiolation in concert with GLK1
Article
Cotyledon de-etiolation is a crucial process during seedling photomorphogenesis that comprises growth pattern reprogramming and the development of photosynthetically active chloroplasts. Studying this process at the molecular level is essential to understand how plants successfully transition to autotrophic metabolism.
We studied the role of the Arabidopsis class II TEOSINTE BRANCHED1, CYCLOIDEA, and PCF (TCP) transcription factor TCP10 in cotyledon de-etiolation using a variety of phenotypic, genetic, and molecular approaches.
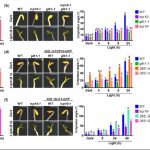
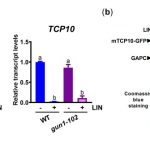
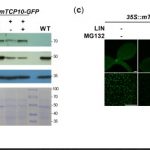
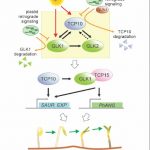
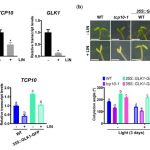
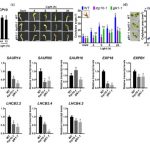
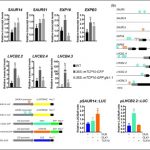
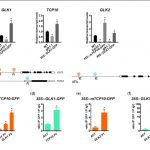
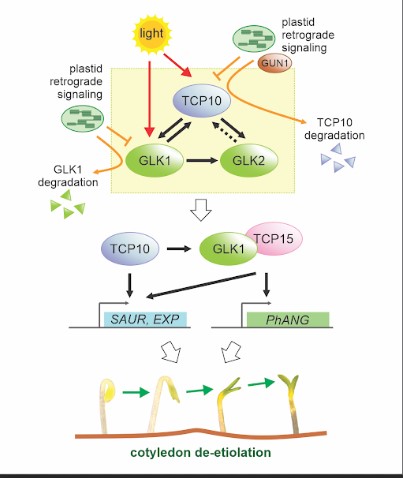
We found that TCP10 is a positive regulator of cotyledon photomorphogenesis that promotes expression of photosynthesis and cotyledon opening genes during de-etiolation, acting either directly or through the central regulator of photomorphogenic development GOLDEN2-LIKE1 (GLK1). Furthermore, TCP10 and GLK1 form a direct positive transcriptional loop that coordinates gene expression responses during de-etiolation. Besides, while light induces TCP10 expression in cotyledons of dark-germinated seedlings to trigger photomorphogenesis, retrograde signals emitted from damaged plastids prevent its expression and induce TCP10 protein degradation via the 26S proteasome, thereby preventing TCP10-induced target gene expression and cotyledon de-etiolation under these conditions.
We propose that the TCP10-GLK1 transcriptional module acts as a hub that integrates light and plastid retrograde signaling pathways to fine-tune de-etiolation and ensure successful early seedling establishment according to internal and environmental stimuli.
{kind=link}